
 Nutrition
et production des carbonates chez les scléractiniaires
Nutrition
et production des carbonates chez les scléractiniaires
par Dietrich Schlichter - DATZ 10, 11, 12/1999 & 1/2000
Bien que les scléractiniaires constructeurs de récifs se nourrissent essentiellement des produits de la photosynthèse provenant de leurs algues symbiotiques, ils disposent de "ressources annexes" qui sont même devenues la source principale chez certaines espèces. *
Le rôle des scléractiniaires dans la croissance du récif
Nombre d'organismes producteurs de calcaire participent à la construction
des récifs récents des biotopes chauds, baignés de
lumière des zones d'eau peu profonde. Parmi ceux-ci, ce sont les
scléractiniaires et les algues calcaires formant des colonies qui
constituent les fournisseurs de calcaire les plus importants. L'importance
des scléractiniaires et des algues calcaires dans la formation
des récifs ne se limite pas seulement à la production de
calcaire. Entre les colonies de coraux se prennent des branches de coraux
brisées, des coquilles d'escargots et de moules ainsi que de minuscules
éléments de squelettes d'unicellulaires, de coraux mous,
d'éponges et d'échinodermes, c'est à dire que ces
débris retenus contribuent également à la croissance
des récifs. En outre les coraux représentent des obstacles
pour le courant et renforcent la sédimentation des particules en
suspension, qui contribuent également par la suite au développement
des récifs. Dans une mesure non moins importante de telles particules
calcaires microscopiques sont produites par des éponges perforatrices,
des oursins et des poissons perroquets.
Afin que ceci aboutisse à la formation de structures récifales
stables et durables, qui résistent aux effets du ressac, le matériau
calcaire d'abord amassé de façon lâche doit être
consolidé. Les scléractiniaires à croissance en forme
de croûte, les bryozoaires, les éponges et surtout les algues
calcaires jouent un rôle dominant ainsi que les microorganismes
sécrétant du calcaire dont l'importance a longtemps été
sous-estimée.
Les processus de cimentation et de solidification contribuent à
la stabilisation ultérieure. Les processus de construction et de
décomposition des récifs sont en très grande partie
le fait d'êtres vivants. L'énergie, néœssitée
pour cela, est acquise par la prise de nourriture. Les interactions entre
alimentation et croissance seront abordées plus loin.
"Animaux photautotrophe"
Les organismes nécessitent de !'énergie en permanence :
1. pour entretenir l'ensemble des processus vitaux
2. pour croître
3. pour se multiplier.
Les plantes sont connues pour pouvoir synthétiser elles-mêmes
les substances organiques nécessaires. La photosynthèse
permet de stocker biochimiquement l'énergie rayonnée. Les
plantes et les cyanobactéries qui possèdent ces propriétés
sont désignées comme photoautotrophe. A l'opposé
les animaux se nourrissent de manière hétérotrophe:
ils dépendent de substances nutritives riches en énergie,
provenant d'autres êtres vivants (de plantes ou d'animaux).
 De
manière intéressante lors du développement d'organismes
sur notre terre les animaux et les plantes se sont associés en
nouvelles combinaisons d'organismes (organismes doubles), qui possèdent
ainsi aussi bien des caractéristiques végétales qu'animales.
Les deux partenaires apportent des contributions propres dans la vie commune
et profitent ainsi des productions du partenaire. Cette forme de vie commune
est appelée symbiose.
De
manière intéressante lors du développement d'organismes
sur notre terre les animaux et les plantes se sont associés en
nouvelles combinaisons d'organismes (organismes doubles), qui possèdent
ainsi aussi bien des caractéristiques végétales qu'animales.
Les deux partenaires apportent des contributions propres dans la vie commune
et profitent ainsi des productions du partenaire. Cette forme de vie commune
est appelée symbiose.
Par l'association, un être vivant inédit avec des caractéristiques
adaptatives élargies est apparu, qui a plus de possibilités
que ne l'avaient les sous-unités correspondantes.
Les scléractiniaires tropicaux et subtropicaux constructeurs de
récifs appartiennent aux organismes doubles de ce type de combinaison,
les plus couronnés de succès au cours de l'évolution.
Par l'insertion de micro-algues dans leurs corps ils sont devenus photoautotrophes
fonctionnels. Cette évolution a été décisive
pour l'approvisionnement énergétique des scléractiniaires
dans des biotopes ayant une offre alimentaire hétérotrophe
réduite (densité d'énergie alimentaire) et ainsi
pour leur succès depuis au moins 200 millions d'années.
Eléments de l'organisation des scléractiniaires
Les scléractiniaires constructeurs de récifs sont soudés
entre eux par un minimum de quelques milliers, animaux individuels de
construction génétiquement identiques (polypes). La construction
de sous-unités ayant des fonctions identiques apporte des avantages.
Ceci se manifeste particulièrement par l'approvisionnement intégré
en substances nutritives à l'intérieur d'une colonie et
en cela, que les colonies réagissent de manière plus flexible
aux modifications de l'environnement. Des surfaces libres ou devenues
libres peuvent être immédiatement colonisées par des
individus nouvellement formés. La formation de colonies rend possible
la conquête de l'espace libre de la colonne d'eau. Les dommages
de tout genre (mécaniques, par des parasites, par des prédateurs)
peuvent être compensés par de nouveaux individus. Ces possibilités
ne sont accessibles aux coraux individuels, existant aussi parallèlement
aux espèces coloniales, que de manière limitée. Le
plan de construction des scléractiniaires est par principe simple
et original, ayant fait ses preuves dans ses traits fondamentaux depuis
550 millions d'année.
La cavité gastrique centrale d'un polype individuel est entourée
d'un cloison corporelle qui est constituée d'une couche externe
et interne de tissus; entre les deux se trouve une couche presque libre
de cellules. On trouve la même structure dans les zones corporelles
entre les polypes. Dans la cloison corporelle externe sont aussi situées,
outre les divers types de cellules, les cellules urticantes (cnidocytes,
nématocystes) , qui ont donné son nom à cet embranchement
(Cnidaria). Elles sont utilisées pour la défense et la capture
des proies. Dans les nombreuses cellules de la cloison corporelle interne
(gastroderme) vivent des micro-algues unicellulaires (diamètres
de 8 à 12 micromètres), qui appartiennent au plan systématique
aux dinoflagellés. Elles se trouvent dans des vacuoles enveloppées
d'une membrane situées dans le cytoplasme des cellules hôtes.
Le nom de micro-algues - zooxanthelles - repose sur leur coloration brun
jaune à brun foncé et sur le fait qu'elles vivent dans les
animaux. La coloration des coraux dépend en partie des zooxanthelles.
Les espèces de coraux possédant des zooxanthelles sont désignées
sous le nom de zooxanthellates, les espèces sans zooxanthelles
comme azooxanthellates (Schumacher & Zibrowius 1985).
A l'aide des méthodes de biologie moléculaire il a été
possible de démontrer que les zooxanthelles vivant dans diverses
espèces de coraux, anémones, moules et unicellulaires ne
font pas partie d'une espèce unique, comme cela était supposé
dans le passé, mais par contre de plusieurs espèces. La
cavité gastrique est subdivisé en septes. Ceci augmente
la stabilité et agrandit la surface interne. La digestion et la
résorption de nourriture s'en trouvent améliorés.
La cavité gastrique est en liaison avec l'eau environnante par
l'intermédiaire d'une ouverture corporelle. Les cavités
gastriques des polypes individuels d'une colonie communiquent entre elles
et forment un système gastrovasculaire, qui a plusieurs fonctions:
1. Dans celui-ci les proies sont prédigérées. La
digestion finale s'opère à l'intérieur des œllules
du gastroderme.
2. La nourriture est répartie à l'intérieur de la
colonie par l'intermédiaire du système gastrovasculaire.
3. L'eau dans système gastrovasculaire soutient l'approvisionnement
en oxygène et l'évacuation des produits finals du métabolisme.
4. La cavité gastrique remplie de liquide joue un rôle important
lors des mouvements de recherche et de capture en tant que "squelette
liquide".
 Des
tentacules se trouvent autour de l'ouverture buccale de la plupart des
espèces de coraux, qui sont essentiellement utilisés lors
de la capture de proies.Le plus souvent l'ensemble de la surface corporelle
est recouverte de cils, qui acheminent des particules de nourriture vers
la bouche ou évacuent des particules de sédiments de la
surface.
Des
tentacules se trouvent autour de l'ouverture buccale de la plupart des
espèces de coraux, qui sont essentiellement utilisés lors
de la capture de proies.Le plus souvent l'ensemble de la surface corporelle
est recouverte de cils, qui acheminent des particules de nourriture vers
la bouche ou évacuent des particules de sédiments de la
surface.
Le squelette calcaire est sécrété en dehors du corps
dans la couche de tissus externe (épiderme) qui lui est consacrée.
Il se compose de carbonate de calcium du type aragonite. Les calices du
squelette sur lesquels se trouvent les polypes se différencient
énormément en taille et forme chez les espèces individuelles.
Il y a des espèces avec des polypes minuscules (un millimètre
de diamètre) comme par exemple les "coraux cornes de cerf'
(Acropora sp.). En outre il y a aussi des espèces à
gros polypes comme les "coraux coupe" (Lobophyllia sp.)
d'un diamètre de plus de 15 cm. La taille des polypes a été
rapprochée par le passé avec celle de la nourriture capturée
; mais une telle corrélation n'existe pas.
Le tissu vivant d'un corail, qui recouvre le squelette d'un film aussi
mince que la peau (0,1 à 0,2 mm) ne représente qu'environ
1 % du poids de la colonie. Dans les cas exceptionnels cette part peut
aussi être plus élevée. Les formes de croissance des
colonies de différentes espèces de coraux se différencient
énormément en taille et forme. L'expression de la forme
individuelle des colonies d'une seule et même espèce peut
être très différente conditionnée par l'emplacement,
ce qui signifie que la forme des colonies n'est pas fixée de manière
définie chez de nombreuses espèces de coraux. Un exemple
est fourni par le corail à calice épineux (Echinopora sp.),
dont les squelettes peuvent être massifs ou en forme de doigts.
Les mouvements et la profondeur de l'eau agissent sur la modification,
où le surfacteur profondeur de l'eau comporte différents
facteurs partiels: irradiation du soleil (intensité lumineuse),
température, pression. En outre l'offre en nourriture planctonique
se modifie également en fonction de la profondeur.
Les coraux avec algues symbiotigues ne constituent pas un cas unique
L'association des micro-algues et des représentants des souches
animales les plus diverses en organismes doubles avec des propriétés
adaptatives élargies ne sont pas une singularité dans les
biotopes aquatiques. L'association de plantes et animaux - et la photoautotrophie
fonctionnelle qui en résulte - a été reprise plusieurs
fois indépendamment l'une de l'autre au cours de l'évolution
des organismes. De telles interdépendances de partenaires sont
particulièrement nombreuses dans l'embranchement des Cnidaires
(coraux, anémones de mer, méduses, hydre verte d'eau douce).
En outre existent aussi des associations de micro-algues avec des unicellulaires,
des vers plats, des mollusques et des nudibranches.
Il existe plus de 1000 espèces de coraux dans le monde
Concurrence dans le récif
Les récifs tropicaux et subtropicaux offrent des biotopes pour
de nombreux organismes, qui se concurrencent tous pour une offre en espace
limitée et pour de maigres ressources alimentaires. Les colonies
de coraux sessiles sont vaincues dans la compétition alimentaire
par les espèces mobiles, car ils sont totalement dépendant
de l'approvisionnement fortuit en nourriture. Les colonies croissent dirigées
dans le courant prédominant; ainsi elles peuvent capturer efficacement
les particules alimentaires présentes dans l'eau. Etant donné
que les coraux ne se nourrissent pas seulement comme les animaux mais
aussi comme les plantes, ils connaissent beaucoup de succès dans
le combat pour la survie malgré le handicap de la sessilité
dans un environnement dans lequel les spécialistes pullulent.
Les diverses espèces de coraux d'un récif ne se trouvent
pas seulement en concurrence avec d'autres organismes, qui ont les mêmes
exigences écologiques, mais surtout aussi entre elles.
De nos jours environ 1000 espèces de coraux sont connues (plus
de 500 espèces fossiles) ; dans le monde environ 550 à 600
espèces participent à la construction de récifs,
dont 500 possèdent des zooxanthelles.
Au cours de l'histoire de leur embranchement les espèces individuelles
de coraux ont développé diverses adaptations de physiologie
alimentaire. Par spécialisation certaines espèces sont capables
de valoriser tels composants alimentaires, que d'autres espèces
n'utilisent pas.
La répartition actuelle des diverses espèces de coraux dans
le récif montre qu'au cours de l'évolution elles occupent
des niches spécifiques, où outre d'autres facteurs écologiques
la nourriture disponible sur place joue un rôle.
Alimentation de coraux zooxanthellates
Pour le maintien de l'ensemble des processus vitaux, donc également
pour les actions qui conduisent à la sécrétion du
squelette calcaire, les scléractiniaires nécessitent de
l'énergie, qu'ils absorbent sous la forme de nourriture. Les scléractiniaires
possesseurs de zooxanthelles, et ce sont, presque sans exception, toutes
les espèces constructriœs de récifs (hermatypes), utilisent
en sus des substances nutritives, que leurs fournissent les zooxanthelles.
Les espèces de scléractiniaires qui participent à
la construction des récifs tropicaux d'eau peu profonde, disposent
d'au moins trois possibilités d'approvisionnement en nourriture.
Nourriture particulaire
Lors de récoltes avec des filets planctoniques ou dans des échantillons,
qui ont été prélevé dans le récif,
se trouve une offre riche en espèces de plancton animal et végétal.
La plus grande partie du zooplancton est représentée par
les petits crustacés et leurs larves ainsi que les larves de mollusques,
d'annélides et de poissons. Des estimations énergétiques
ont fourni des indices selon lesquels l'offre en zooplancton de filet
ne suffit pas pour nourrir l'ensemble des coraux, ceci signifie que la
densité d'aliments énergétiques est trop faible.
Des particules organiques mortes de taille les plus diverses sont également
disponibles et proviennent de plantes et d'animaux en décomposition,
d'excréments ou de carapaces provenant de mues de crustacés.
 Outre
le zooplancton relativement grand et les particules organiques il existe
un nombre important de plus petits organismes et de particules organiques
en suspension. Ces composantes alimentaires n'ont pas été
remarquées aux cours des examens passés pour des raisons
de méthodes. La taille des mailles des filets à plancton
ou le diamètre des pores était trop grand et suite à
cela leur signification d'alimentation physiologique ne pouvait être
prise en compte. Des examens réalisés au cours des deux
dernières décennies ont multiplié les informations
selon lesquelles aussi bien les unicellulaires et les bactéries
que les plus petites particules en suspension étaient d'une importanœ
significative en ce qui concerne la structure alimentaire (Sorokin 1993).
Toutes ces composantes sont de nature organique et ainsi par principe
utilisable comme nourriture par les scléractiniaires. Des particules
minérales en suspension sont par contre inutilisables pour l'alimentation
des scléractiniaires.
Outre
le zooplancton relativement grand et les particules organiques il existe
un nombre important de plus petits organismes et de particules organiques
en suspension. Ces composantes alimentaires n'ont pas été
remarquées aux cours des examens passés pour des raisons
de méthodes. La taille des mailles des filets à plancton
ou le diamètre des pores était trop grand et suite à
cela leur signification d'alimentation physiologique ne pouvait être
prise en compte. Des examens réalisés au cours des deux
dernières décennies ont multiplié les informations
selon lesquelles aussi bien les unicellulaires et les bactéries
que les plus petites particules en suspension étaient d'une importanœ
significative en ce qui concerne la structure alimentaire (Sorokin 1993).
Toutes ces composantes sont de nature organique et ainsi par principe
utilisable comme nourriture par les scléractiniaires. Des particules
minérales en suspension sont par contre inutilisables pour l'alimentation
des scléractiniaires.
La concentration des particules en suspension doit être très
faible pour la croissance d'un corail et d'un récif sain. Si la
densité de particules est trop élevée, il se forme
sur les coraux une couche de sédiments dommageable, qui peut conduire
à la mort des colonies. Des colonies en formes de branches sont
moins touchées par la sédimentation que les foliaires (en
forme de feuilles) ou les massives. La plupart des espèces de scléractiniaires
disposent de mécanismes de nettoyage efficaces: par absorption
d'eau dans le système gastrovasculaire le tissu peut être
gonflé, si bien que les particules glissent sur la surface. De
nombreuses espèces éliminent les particules sédimentées
de la surface à l'aide de cils et/ou du mucus.
Les tentacules des scléractiniaires sont bardés de cnidoblastes
remplis de poison. Ceci permet d'en tirer la conclusion qu'ils capturent
et valorisent essentiellement du zooplancton. Effectivement tous les animaux
n'en sont pas capables.
La nourriture particulaire se compose des composantes les plus diverses.
Parmi les organismes planctoniques se trouve de petits crustacés,
des larves de mollusques, d'annélidés, etc. Si le plancton
entre par hasard en contact avec les coraux, il est capturé à
l'aide de toxines, qui sont stockées dans les cnidoblastes. Après
l'injection du poison - l'éjection des cystes s'effectue en l'espace
d'un millième de seconde - la proie se trouve paralysée
et peut être amenée vers la bouche avec l'aide des tentacules
et de l'action des cils puis avalée.
La plupart des espèces de scléractiniaires capturent leurs
proies durant la nuit. Ainsi la probabilité d'acquisition des proies
s'en trouve augmentée.
1. A ce moment les chasseurs actifs de jour, qui chassent "visuellemenf',
sont inactifs et ne soufflent pas leur proie aux coraux.
2. Les proies vivantes ne peuvent pas s'orienter et ne peuvent éviter
l'appareil de capture des coraux.
3. Au cours de la nuit l'offre en zooplancton est nettement plus importante,
étant donné que le plancton ne quitte ses cachettes qu'à
la tombée de l'obscurité.
4. Durant la journée les polypes de la plupart des coraux se retirent
dans les calices protecteurs du squelette, afin d'échapper à
des prédateurs (poissons-perroquets, étoiles de mer, etc.).
Les microorganismes et les particules en suspension: en ce qui concerne
ces composantes alimentaires il s'agit de bactéries, d'unicellulaires
(flagellés, foraminifères, ciliés) et de minuscules
particules organiques. Souvent ce matériau organique mort (la partie
en cellulose) ne peut pas être utilisée directement car les
enzymes de digestion nécessaires font défaut. Les microorganismes
qui poussent sur les particules organiques et s'y multiplient sont digérés
à leur place. Le matériau particulaire organique de taille
microscopique, en suspension (vivant, mort) peut être capturé
par les coraux par l'intermédiaire du mucus. Cette méthode
d'alimentation ne devient significative seulement par l'enrichissement
du mucus en particules.
Substances nutritives dissoutes
Dans l'eau de mer il est possible de détecter de nombreuses substances
organiques, entre autres des glucides, des acides aminés et des
acides gras. Cette matière organique dissoute, rapportée
à un litre d'eau de mer, ne se trouve qu'en concentration extrêmement
faible (quelques millièmes de grammes par litre). L'importante
dissolution signifie que la source de nourriture ne peut être utilisée
que s'il existe des mécanismes pour l'extraction de l'eau et l'accumulation
dans le corps. Grâce à des systèmes de transport spécifiques.
localisés dans les surfaces corporelles externes, les coraux sont
capables d'intégrer ces substances et de les exploiter comme sources
d'énergie ou pour la croissance.
En ce qui concerne les systèmes de transports il s'agit de pompes,
qui transportent dans les cellules les molécules provenant de l'eau
de mer vers des gradients de concentration, ce pour quoi il faut de l'énergie.
Les mécanismes d'intégration pour les acides aminés
ou les glucides chez les coraux sont comparables à ceux du corps
humain, avec l'aide desquels les glucides et les acides aminés
sont transportés des intestins vers le sang.
L'admission des glucides et des acides aminés provenant de l'eau
de mer par les scléractiniaires ne peut pas être analysée
directement dans le récif pour des motifs de méthode. C'est
pourquoi il faut mesurer l'admission de ces substances dans le laboratoire,
afin de pouvoir juger leur signification d'alimentation physiologique.
Si on prend pour base de calcul énergétique les taux d'admission
acquis de manière expérimentale (quantité de substance
résorbée par période) en glucides, acides gras et
acides aminés, il apparaît qu'avec les substances reçues
plus de 30 % du métabolisme de base peut être couvert (Schlichter
1982, 1983).
De la fonction des zooxanthelles et des algues perforatrices, d'un scléractiniaire, qui vit en association avec des algues symbiotiques à 145 mètres de profondeur, et d'une espèce, spécialisée sur les particules les plus fines... *
Les zooxanthelles vivent dans des cellules du gastroderme. Leur nombre par centimètre carré de surface de corail varie énormément (de quelques 100.000 à 10 millions) et dépend de l'espèce de corail et de l'intensité du rayonnement.
Le rôle des algues symbiotigues
Entre les zooxanthelles et le tissu animal il existe des courants de matière
et d'énergie. Les productions de l'hôte et des algues se
complètent de manière idéale. La respiration et la
photosynthèse sont court-circuités sur le plan cellulaire.
Les substances inorganiques (gaz carbonique, combinaisons phosphorées
et azotée) qui échoient comme sous-produits dans le métabolisme
de l'hôte (bien sûr les algues produisent elles-mêmes
également du gaz carbonique lors de la respiration), peuvent immédiatement
être utilisées par les algues pour la synthèse de
combinaisons organiques, ce qui signifie que la dégradation de
combinaisons organiques carbonées productrice d'énergie
et la nouvelle synthèse de combinaisons organiques mue par l'énergie
solaire se déroulent pratiquement sans pertes à l'intérieur
de la cellule hôte. La découverte de ces relations a été
rendue possible par l'utilisation de substances avec marqueur radioactif
(14C). Les processus interactifs représentent un efficace
système de recyclage.
Cette chaîne alimentaire intracellulaire diffère complètement
des chaînes alimentaires traditionnelles. Les maillons individuels
des chaînes alimentaires aquatiques traditionnelles sont séparés
l'un de l'autre dans l'espace et dans la fonction ; le phytoplancton photoautotrophe
(les producteurs primaires) est utilisé par les animaux consommateurs
de végétaux (herbivores), qui de leur côté
constituent de la nourriture pour les consommateurs secondaires. Chez
le corail, être vivant double, ce n'est pas le cas. Les producteurs
primaires - les zooxanthelles - et le consommateur primaire - l'hôte
-, qui est de plus en même temps consommateur secondaire, forment
un combinat de production avec des avantages importants: des substances
nutritives minérales restent à l'intérieur des cellules
hôtes contenant des zooxanthelles et les pertes en "destructeurs"
("minéralisateurs"), qui constituent normalement des
éléments des chaînes alimentaires, cessent complètement.
Si suffisamment d'énergie de rayonnement est disponible, les algues
synthétisent bien plus de combinaisons organiques, qu'elles n'en
ont besoin. La quantité stockée de photoassimilats peut
être si importante que les coraux peuvent en couvrir l'ensemble
de leurs besoins énergétiques. Les coraux n'utilisent les
algues qui les nourrissent que dans un but fonctionnel ; les algues elles
mêmes ne sont pas digérées. Les coraux n'exploitent
que les substances organiques, que les algues dégagent des membranes
des vacuoles vers le plasma cellulaire. Il s'agit de glycerol, de glucose,
d'acides aminés et d'acides gras ; peut-être que les algues
éclusent des lipides par exocytose, c'est à dire emballé
dans des vésicules, dans le cytoplasme de l'hôte. Au sens
figuré les coraux utilisent les producteurs primaires qui vivent
en eux avec le plus de ménagement, en ce sens qu'ils utilisent
seulement le "lait" de la vache, mais qu'ils ne tuent pas la
vache.
L'analyse de coraux fossiles a fourni des indications selon lesquelles
les coraux et les zooxanthelles se sont associés il y a 200 millions
d'années (Stanley & Swart 1995). Durant cette longue coévolution
des adaptations ont eu lieu qui ont rendu effectif la cohabitation d'une
part ; d'autre part des dépendances sont apparues. Si on sépare
les deux partenaires l'un de l'autre, il est possible d'élever
les zooxanthelles c'est à dire que sans hôtes elles sont
encore capables de vivre. Des hôtes sans zooxanthelles vivotent
ou meurent (Yonge 1973). Les hôtes ont donc dû perdre des
capacités physiologiques spécifiques au cours de la coévolution
avec les algues. Comparé avec des algues unicellulaires seules
les zooxanthelles vivent, en ce qui concerne leur approvisionnement en
substances nutritives minérales, dans des conditions plus favorables
et il faut s'attendre à ce qu'elles se multiplient mieux dans l'hôte
que dans une culture. Ce qu'il est possible de mesurer est que la productivité
des zooxanthelles est plus importante dans l'hôte que dans des conditions
de culture et que les algues présentes dans l'hôte contre
toute attente ont un taux de multiplication plus faible. La production
primaire ne rentre donc pas dans la multiplication des zooxanthelles mais
profite à l'hôte. Ceci signifie que l'hôte contrôle
le métabolisme des symbiontes et l'exportation de photoassimilat
dans le plasma de la cellule.
En ce qui concerne les facteurs de l'hôte il s'agit, comme des recherches
récentes l'on montré, d'un mélange d'acides aminés,
qui augment la perméabilité des membranes des vacuoles pour
les photoasimilats (Gates et al. 1995). La population des algues symbiotiques
(pas seulement les individuelles) "se paye" par un taux de multiplication
nettement plus faible de la protection conservatrice de l'espèce
dans l'hôte armé de cnidoblastes.
Les zooxanthelles des cellules hôtes se multiplient par division.
Malgré le contrôle de la division par l'hôte la capacité
de réception en algues des cellules hôtes a été
dépassée. Les algues excédentaires sont exclues du
cytoplasme et relâchées dans la mer. Le fait que les zooxanthelles
en excédent tout comme les plus anciennes ne soient pas digérées
par les coraux - l'émission dans la mer est un luxe d'un point
de vue énergétique - est encore largement inconnu.
 Pourquoi
justement les coraux ont ils choisi des dinoflagellés comme partenaires
et pas des petites algues unicellulaires vertes ou des cyanobactéries,
qui apparaissent comme symbiontes dans d'autres embranchements (unicellulaires,
éponges, ciliés) ? Une importante condition pourrait être
qu'aussi bien le métabolismes des zoxanthelles que celui de l'hôte
animal soit basé sur les lipides et que les deux métabolismes
puissent se court-circuiter biochimiquement sans problème pour
cette raison, ceci signifie que la compréhension biochimique était
sans problème, étant donné que les deux partenaires
"parlent la même langue". Un métabolisme cellulaire,
dans lequel les lipides ont une signification centrale, est inhabituel
chez des algues unicellulaires, mais distingue les dinoflagellés.
Pourquoi
justement les coraux ont ils choisi des dinoflagellés comme partenaires
et pas des petites algues unicellulaires vertes ou des cyanobactéries,
qui apparaissent comme symbiontes dans d'autres embranchements (unicellulaires,
éponges, ciliés) ? Une importante condition pourrait être
qu'aussi bien le métabolismes des zoxanthelles que celui de l'hôte
animal soit basé sur les lipides et que les deux métabolismes
puissent se court-circuiter biochimiquement sans problème pour
cette raison, ceci signifie que la compréhension biochimique était
sans problème, étant donné que les deux partenaires
"parlent la même langue". Un métabolisme cellulaire,
dans lequel les lipides ont une signification centrale, est inhabituel
chez des algues unicellulaires, mais distingue les dinoflagellés.
Outre les substances nutritives, que les zooxanthelles fournissent à
leur hôte, leur approvisionnement en oxygène est aussi considéré
comme production symbiotique. Une importante production d'oxygène
photosynthétique ne semble en aucun cas être un avantage.
Au contraire des concentrations en oxygène élevées
(sous forme de liaisons agressives) ont un effet dommageable sur l'hôte
et il possède des enzymes spéciales afin d'éliminer
les combinaisons oxygénées toxiques.
Le déficit en oxygène est-il compensé par les
zooxanthelles ?
Des recherches plus récentes confirment que les zooxanthelles contribuent
en présence d'un mouvement d'eau faible à une compensation
plus rapide du déficit en oxygène survenu durant la nuit.
Comme toutefois l'eau est toujours en mouvement sur le récifs cet
aspect pourrait avoir une signification subalterne.
Le déploiement nocturne des polypes, discuté avant dans
le contexte de la capture de proies, peut aussi être mis de manière
spéculative en relation avec l'approvisionnement en oxygène.
Durant la journée il y a une telle production d'oxygène
due à la photosynthèse des algues, qu'il n'y a pas besoin
de le prendre dans l'eau environnante ; au contraire, l'oxygène
diffuse à partir du tissu des coraux dans l'eau. Durant la nuit
par contre l'oxygène doit diffuser vers le tissu à partir
de l'eau, étant donné que durant l'obscurité les
coraux et les algues respirent de l'oxygène. Lorsque la nuit les
polypes sont ouverts et sont baignés par l'eau riche en oxygène,
l'approvisionnement en oxygène est facilité par l'intermédiaire
de la diffusion. Dans l'eau riche en oxygène des récifs
l'approvisionnement en oxygène ne semble pas constituer un problème
fondamental, surtout lorsqu'on se souvient que les distances de diffusion
dans les tissus des coraux atteignent un maximum de 100 à 200 micromètres.
Algues endolithiques - une autre source de nourriture
Outre les zooxanthelles les scléractiniaires sont capables d'utiliser
une autre source de nourriture végétale, c'est à
dire photoautotrophe: dans le squelette de très nombreuses espèces,
soit avec soit sans zoxanthelles, vivent des algues vertes et des algues
rouges filamenteuses (Osreobium quekettii, Porphyra sp.). Entre
ces algues perforatrices et le tissu des coraux des échanges de
matière, comparables à celui tel qu'il a été
décrit pour les zoxanthelles. La preuve d'un courant de matière
a réussi par l'utilisation de carbone radioactif (Schlichter et
al. 1995). Au moyen de ce marquage des substances provenant des algues
ont pu être rendues visibles dans les tissus des coraux.
La contribution de la symbiose des algue endolithiques au métabolisme
total est nettement plus faible que celle des zooxanthelles. Ceci est
certainement en rapport avec le fait que le métabolisme des cellules
des algues filamenteuses n'est pas aussi étroitement lié
avec l'hôte. En outre le métabolisme des algues vertes constitue
un métabolisme glucidique typique et pas un métabolisme
lipidique, comme cela est caractéristique pour les zooxanthelles.
Les membranes des deux partenaires se touchent seulement c'est à
dire que les courants de matière ne sont pas court-circuités
à l'intérieur d'une cellule. Il y a des situation qui sont
comparables avec les bases structurelles et fonctionnelles chez les lichens.
Dans cette symbiose des champignons (hétérotrophes) et des
algues vertes unicellulaires se sont associés.
Les conditions de faible éclairage dans le squelette couvert de
tissu ne permettent qu'une faible production primaire des algues perforatrices,
qui représente environ 7 % de la productivité des zooxanthelles.
Même si l'apport énergétique au métabolisme
des coraux par les algues perforatrices est relativement réduit,
elles ont cependant de l'importance dans le contexte de la conservation
des substances nutritives à !'intérieur de l'écosystème
récifal et leur fonction écologique est comparable avec
celle des zoxanthelles (Schlichter et al. 1995, 1996). L'utilisation de
photoassimilats d'algues endolitihiques peut constituer une source complémentaire
de nourriture surtout pour les algues sans zoxanthelles. Avec l'énergie
ainsi mise à disposition des goulots d'étranglement dans
l'approvisionnement en nourriture particulaire peuvent être mieux
surmontés.
Sources de nourriture et besoins énergétiques de la
symbiose
Grâce aux différentes possibilités d'alimentation
l'approvisionnement énergétique des coraux zooxanthellates
est stable. Une source de nourriture, qui amène momentanément
peu d'énergie au corps, par exemple le zooplancton, peut être
compensée par une autre. Dans un cas isolé l'optimisation
de stratégies d'alimentation peut même complètement
permettre de se passer des autres. Ainsi il existe une espèce de
corail mou (Heteroxenia fuscescens), qui ne capture aucun zooplancton
et aucune particule organique et qui se nourrit exclusivement par l'absorption
de composés organiques dissous et des produits de la photosynthèse
des zooxanthelles (Schlichter 1982, 1983).
L'importance de l'apport énergétique, par lequel chaque
possibilité d'alimentation contribue au métabolisme de base
de l'organisme double, ne peut pour l'instant qu'être estimé,
étant donné qu'il est très difficile sur le plan
de la méthode, de mesurer avec exactitude l'ensemble des possibilités
d'alimentation d'une seule et même espèce. L'examen de 23
espèces de scléractiniaires de la Grande Barrière
a fourni des éléments d'évaluation, dont la validité
dépend toutefois en grande partie de la disponibilité en
lumière de l'habitat: 20 % par exploitation de nourriture particulaire,
10 % par absorption de composés organiques dissous et 70 % par
utilisation des assimilats des algues symbiotiques (Sorokin 1993).
Nourriture particulaire microscopique
L'offre en nourriture particulaire d'un récif est diverse et s'étend
des bactéries au plancton plus grand; outre des particules organiques
de taille les plus diverses en font partie. La quantité correspondante
des composants individuels (densité de nourriture énergétique)
dépend fortement des possibilités locales du récif,
de l'exposition au vent et des conditions hydrographiques qui en résultent.
L'élément central de nos recherches pluriannuelles dans
le golfe d'Akaba a été de découvrir les adaptations
physiologiques des coraux durs, qui leurs permettent d'utiliser certains
composants de l'offre alimentaire particulaire. Comme hypothèse
de travail nous avons posé la question: y a t'il des corrélations
entre l'emplacement, la disponibilité de nourriture en fonction
de la profondeur et les stratégies de capture des proies (stratégies
d'acquisitÎon de nourriture) des scléractiniaires ?
L'offre de nourriture se modifie quantitativement et qualitativement en
fonction de la profondeur. La production primaire des algues planctoniques
a lieu dans les couches supérieures de l'eau en corrélation
avec une offre supérieure en zooplancton. L'offre en zooplancton
diminue avec l'augmentation de la profondeur et la signification de la
physiologie alimentaire des unicellulaires, bactéries et matériel
organique mort sombrant, augmente à l'opposé. Parmi les
scléractiniaires certaines espèces utilisent des composants
très précis de l'offre hétérogène en
nourriture particulaire. Elles se sont spécialisées dans
la capture des particules les plus petites, qui ne sont pas attirantes
pour des chasseurs actifs. Dans le cas de ces composants alimentaires
il s'agit de bactéries et d'unicellulaires ou de minuscules particules
organiques. La dépense énergétique et temporelle
pour la capture de telles proies, est importante d'une part pour les chasseurs
et le gain énergétique minimal d'autre part. Le problème,
de retenir et de concentrer les particules en suspension, est solutionné
de manière très différente par les espèces
individuelles de coraux et les mécanismes de capture et de concentration
reposent sur des principes complètement divers.
Le corail dur à zooxanthelles Leptoseris fragilis vit par
des profondeurs comprises entre 90 et 145 mètres, donc loin de
la couche supérieure productive et riche en plancton. Les coraux
en forme de soucoupes dépourvus de tentacules atteignent un diamètre
pouvant atteindre huit centimètres. Ils orientent la partie corporelle
pourvue de la bouche vers le haut. Des particules sombrant sont capturées
par un piège à sédiments (collecteur) et menées
vers la bouche le long des chenaux entre les septes calcaires à
l'aide des cils. Une abondante garniture de cils dans la gorge transporte
les particules vers les cavités gastriques fortement ramifiées,
où elles restent retenues comme par un filtre. Cela avec les particules
apportées par l'eau ne quitte pas le corps par l'ouverture buccale
comme c'est la règles chez les autres espèces de cnidaires,
mais par de minuscules pores de la thèque externe. Le système
de "traversée", au cours duquel des particules sont retenues
dans un fin système gastrovasculaire ramifié à partir
d'un courant auto généré, provoque une concentration
effective dans la cavité gastrique, ce qui améliore la situation
alimentaire. Etant donné qu'il y a peu de courant dans l'espace
vital de cette espèce de corail, seul le courant autogénéré
peut conduire à une concentration de particules.
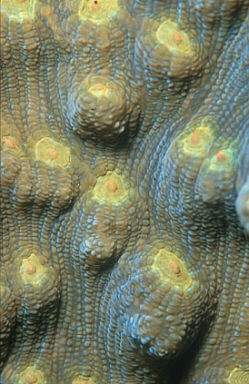 L'espèce
de scléractiniaire également sans tentacules Mycedium
elephantotus, qui est particulièrement abondante à des
profondeur comprises entre 20 et 40 mètres, est un pêcheur
à filet de mucus. Les colonies de cette espèce sont construites
à base de plaques de squelettes en forme d'éventail, qui
se dressent obliquement dans l'eau. Les plaques constituent des obstacles
au courant et fonctionnent comme des attrape mouches. Le tissu extérieur
(Epiderme) contient d'innombrables cellules de mucus, la surface est couverte
d'une épaisse couche de mucus et les cils sont absents. Des particules
en suspension dans l'eau effleurent la surface et sont enveloppées
de mucus à la vitesse de l'éclair (en l'espace d'une seconde).
S'il s'agit de particules minérales, elles descendent des plaques
en glissant. Des particules organiques (dans l'expérience de la
nourriture de poisson pulvérisée a été utilisée)
sont capturées avec le "filet" de mucus. Les petits fils
du filet s'emmagasinent en éléments plus gros ayant pour
résultat une concentration de particules alimentaires. Les agrégats
contenant des particules alimentaires glissent le long des plaques. Elles
touchent fortuitement une ou plusieurs ouvertures buccales et sont avalées
(Schlichter & Brendelberger 1998).
L'espèce
de scléractiniaire également sans tentacules Mycedium
elephantotus, qui est particulièrement abondante à des
profondeur comprises entre 20 et 40 mètres, est un pêcheur
à filet de mucus. Les colonies de cette espèce sont construites
à base de plaques de squelettes en forme d'éventail, qui
se dressent obliquement dans l'eau. Les plaques constituent des obstacles
au courant et fonctionnent comme des attrape mouches. Le tissu extérieur
(Epiderme) contient d'innombrables cellules de mucus, la surface est couverte
d'une épaisse couche de mucus et les cils sont absents. Des particules
en suspension dans l'eau effleurent la surface et sont enveloppées
de mucus à la vitesse de l'éclair (en l'espace d'une seconde).
S'il s'agit de particules minérales, elles descendent des plaques
en glissant. Des particules organiques (dans l'expérience de la
nourriture de poisson pulvérisée a été utilisée)
sont capturées avec le "filet" de mucus. Les petits fils
du filet s'emmagasinent en éléments plus gros ayant pour
résultat une concentration de particules alimentaires. Les agrégats
contenant des particules alimentaires glissent le long des plaques. Elles
touchent fortuitement une ou plusieurs ouvertures buccales et sont avalées
(Schlichter & Brendelberger 1998).
Le transport le long de la surface du corail a lieu par gravitation; un
faible courant a un effet de soutien. En présence d'un mouvement
d'eau plus puissant les fils de mucus chargés de particules sont
arrachés de la surface et sont perdus en tant que nourriture. Afin
que cela ne se produise pas et que cette forme de récolte de nourriture
soit efficace, ainsi peut on argumenter, des lieux ayant une énergie
en eau élevée n'entrent pas en ligne de compte comme espace
vital. Des conditions favorables règnent à partir de profondeurs
de 15 mètres. La distribution principale de M. elephantotus
se situe entre 20 et 30 mètres; l'espèce est cependant encore
abondante par 40 mètres et atteint à cette profondeur à
Akaba une couverture vitale du récif de l'ordre de 25 %.
Une offre accrue en matière en suspension est responsable de la
présence de Mycedium elephantotus, outre une pauvreté
de courant dans l'espace vital. L'offre en particules plus importante
est due au fait que par des profondeurs de 15 à 20 mètres
les vagues superficielles (orbitales) et les courants profonds laminaire
se rencontrent. (Riedl 1969 ; Roberts et al. 1975). Les filets de mucus
concentrés en particules arrachées de la surface des coraux
par des mouvements d'eau plus puissants sont de nouveau utilisés
comme nourriture par les crustacés et les poissons (Benson &
Muscatine 1974).
Le transport de la nourriture enrobée de mucus vers la cavité
gastrique est effectué par les cils. Le pharinx possède
une couverture de cils très dense avec l'aide desquels la "saucisse
de mucus" est transportée comme sur un tapis roulant vers
la cavité gastrique. L'absorption de nourriture se déroule
en partie très "violemment" et sans coordination: diverses
bouches cherchent en même temps à avaler la même "saucisse
de mucus".
Certains scléractiniaires disposent de mécanismes d'amplification de la lumière pour l'effectivité du rendement de la photosynthèse de leurs zooxanthelles. *
Les fondements de l'association des zooxanthelles et des coraux ont déjà été présentés, de même que le recyclage intracellulaire des substances nutritives.
Augmentation de la photosynthèse en présence de lumière
faible
Le niveau de la valeur énergétique, à laquelle les
algues contribuent pour le métabolisme de leur hôte, est
influencé par divers facteurs. Trois d'entre eux doivent être
traités à titre d'exemple:
1. La quantité de photoassimilats transférés dépend
du nombre d'algues symbiotiques présent dans les tissus de l'hôte.
La production photosynthétique des zooxanthelles est d'autre part
influencée par la température, la salinité et l'intensité
du rayonnement. Le rayonnement solaire direct des eaux tropicales et subtropicales
sous la surface est souvent si puissant que les algues ou des coraux entiers
sont endommagés (aussi par la quantité élevée
en UV-B/C). C'est pourquoi la croissance n'est pas optimale dans les zones
récifales peu profondes. Le facteur d'irradiation, avec les anomalies
de température semble être responsable pour le blanchiment
(bleaching), au cours duquel les hôtes perdent leurs zooxanthelles.
Ce processus de blanchiment entraîne la mort des coraux et l'arrêt
de la croissanœ du récif.
2. Un autre facteur extérieur qui influence la photosynthèse
des algues vivant en liberté et les zooxanthelles, est l'offre
en substances nutritives minérales. Dans les eaux oligotrophes,
dans lesquelles les coraux prospèrent, leur concentration et très
faible si bien que la production primaire des algues est limitée
par carence en sels nutritifs (azote, phosphates, fer), bien que l'énergie
de rayonnement soit disponible à volonté. L'approvisionnement
des zooxanthelles en substances nutritives minérales est meilleure
car ils profitent des produits finals du métabolisme de leur hôte.
3. Le niveau des besoins énergétiques des zooxanthelles
par rapport au métabolisme de l'hôte est nettement déterminé
par le besoin énergétique correspondant de l'espèce
du corail hôte. S'il est réduit, les photoassimilats représentent
une proportion importante. Si le besoin énergétique de l'hôte
est élevé, avec une productivité identique des zoxanthelles,
la proportion a moins d'importance.
© Extrait de la Gazette Marine N° 75 avril
2003 et 76 - juillet 2003
Publication réservée aux membres des Amis de l'Aquarium
1932.